








摘要
猪流行性腹泻病毒(PEDV)是影响全球养猪业的主要病原体,其暴发会造成重大经济损失。自1970年被发现以来,PEDV的遗传变异性给疫苗研发带来了诸多挑战,这一问题在2010年PEDV全球流行后变得更为严峻。由于现有疫苗对PEDV毒株的交叉保护有限,且多价PEDV疫苗的研发仍探索不足阶段,因此迫切需要一个疫苗改进方案。严重急性呼吸综合征冠状病毒2型(SARS-CoV-2)疫苗的快速研发以及正在进行的泛沙贝科病毒疫苗研究,已证明新一代疫苗平台和新型抗原设计策略的潜力。这些研发进展为开发多联PEDV疫苗提供了宝贵的经验。本综述总结了PEDV病毒学方面的关键内容,并结合SARS-CoV-2疫苗的创新探索多联疫苗开发,提出了一个开发新型PEDV疫苗解决方案的框架。
关键词
猪流行性腹泻病毒(PEDV)、SARS-CoV-2、冠状病毒、多价疫苗、广谱保护疫苗、抗原设计、疫苗平台、刺突蛋白(S蛋白)、中和抗体、新一代疫苗
1、引言
1.1 PEDV的全球影响与多联疫苗的需求
猪流行性腹泻病毒(PEDV)是一种冠状病毒,可引起猪流行性腹泻(PED),是一种传染性极强的肠道疾病,在易感猪群中可导致1至3日龄仔猪近100%的死亡率。虽然较大日龄猪只死亡率较低,但PEDV感染仍显著影响生长性能,造成经济损失。怀孕后备母猪和经产母猪感染PEDV后会损害繁殖性能,导致分娩率降低、返情和流产增加、木乃伊胎率升高,给养猪业带来的巨大流经济损失。
20世纪70年代早期,PEDV首次英国被发现,高致病性PEDV毒株在2010出现于中国,成为中国仅次于猪繁殖与呼吸综合征病毒(PRRSV)的第二大重要传染性病原体。在全球范围内,PED仍是仔猪死亡的主要原因。PEDV于2013年在北美被开始流行,导致美国一年内损失了10%的猪只,造成严重损失。2013-2014年的PED疫情导致美国经济损失达9亿至18亿美元,这凸显了养猪业对PED流行有效预防策略的迫切需求。
2014年,PEDV传播至加拿大,安大略省的首次暴发与受污染的饲料有关,随后传播到魁北克省和马尼托巴省的农场。加拿大借鉴了美国的经验教训,通过严格的生物安全措施有效控制了该病毒。然而,该病毒尚未在北美洲根除,其持续的威胁强调了将疫苗研发作为预防策略的关键组成部分的必要性。
1.2 PEDV与SARS-CoV-2疫苗研发面临的共同挑战与策略
作为冠状病毒科的成员,PEDV和严重急性呼吸综合征冠状病毒2型(SARS-CoV-2)的疫苗研发都面临着相似挑战,这主要源于抗原变异和免疫逃逸策略。这两种病毒的刺突蛋白(S蛋白)—中和抗体的主要靶点—频繁发生突变,使得开发广谱保护性疫苗非常复杂。PEDV存在两个基因组群和至少五个亚型,与SARS-CoV-2出现的Alpha、Delta和Omicron等变种类似,这些变种降低了疫苗效力。此外,PEDV和SARS-CoV-2均具有高度的基因重组能力,这使得免疫逃逸成为可能,并可能导致更强毒力毒株出现。
在2019冠状病毒病(COVID-19)大流行期间,SARS-CoV-2疫苗的快速研发标志着疫苗学的重大突破,将传统疫苗开发周期(人用疫苗10-15年,兽用疫苗3-6年)压缩至不到一年。这一前所未有的进展得益于对SARS-CoV和MERS-CoV的前期研究、临床试验阶段的重叠、紧急资金支持,以及在获得监管批准前承担财务风险启动生产。这些研究充分展现了重组蛋白、mRNA和病毒载体平台的适应性,以及基于结构和嵌合抗原设计策略的潜力,即既能精准靶向保守病毒表位,又能快速应对新出现的变异株。这些突破性进展凸显了PEDV疫苗应该超越第一代灭活和减毒疫苗的重要性。通过整合新一代平台与合理抗原设计,能更有效地应对不断演变的病毒威胁。
本综述探讨了针对SARS-CoV-2疫苗策略(尤其是泛沙贝病毒保护策略)如何指导开发有效的多价PEDV疫苗。通过融合PEDV病毒学与SARS-CoV-2疫苗研发的成果,本研究旨在推动改良疫苗的研发,使其能够为养猪业提供更广泛、更具交叉保护性的免疫效果。
2、PEDV病毒学、致病机制与免疫应答
2.1病毒学与致病机制
PEDV属于冠状病毒科(Coronaviridae)、α冠状病毒属(Alphacoronavirus),它与SARS-CoV-2及其他冠状病毒具有相似结构。值得注意的是,冠状病毒拥有RNA病毒中已知的最大基因组,这导致了其遗传多样性。
2.1.1基因组与病毒粒子结构
PEDV基因组为线性单股正链RNA,大小约为28 kb,基因组包含5'端帽结构和3'端poly-A尾,两端侧翼为非翻译区(UTRs)。基因组按顺序编码七个开放阅读框(ORFs):ORF1a、ORF1b、S、ORF3、囊膜蛋白(E)、膜蛋白(M)和核衣壳蛋白(N)。结构蛋白ORFs(S、E、M、N)和辅助蛋白ORF3通过一个不连续的过程转录,产生一套嵌套的亚基因组mRNAs(sgmRNAs)。相比之下,基因组RNA直接作为模板翻译ORF1a和ORF1b成多聚蛋白pp1a和pp1ab,随后被病毒蛋白酶加工成16种非结构蛋白(nsp1-nsp16)。这些nsps执行基本功能,例如形成复制-转录复合体(RTC)和逃避先天免疫应答。尽管RTC具有校对能力,PEDV和其他冠状病毒基因组仍会积累突变,导致新毒株的出现。
2.1.2病毒粒子结构与结构蛋白
PEDV病毒粒子是有包膜的球形颗粒,直径范围为90至160 nm。与SARS-CoV-2中观察到的一样,其最显著的特征是由多个刺突蛋白(S蛋白)拷贝形成的突出“冠冕”(12-15 nm)。S蛋白是病毒表面最丰富的蛋白,除S蛋白外,PEDV病毒粒子还包含其他三种主要结构蛋白:膜蛋白(M)、囊膜蛋白(E)和核衣壳蛋白(N)。
S蛋白是同源三聚体的I类融合糖蛋白,对病毒附着宿主细胞及随后的进入至关重要,每个单体是一个分子量为160-220 kDa的跨膜蛋白,由两个不同的亚基组成:介导受体结合的S1和促进膜融合的S2。由于其免疫原性,S1亚基是中和抗体和疫苗开发的主要靶标。M蛋白是一种保守的20-30 kDa跨膜糖蛋白,是最丰富的结构蛋白,与S、E和N蛋白的相互作用,对包膜稳定性和病毒组装至关重要。E蛋白是一种8-12 kDa的跨膜糖蛋白,通过与M蛋白和其他病毒成分的相互作用,促进病毒组装、出芽和释放。N蛋白是一种磷酸化的约58 kDa蛋白,包裹病毒RNA基因组形成核糖核蛋白复合体,并在病毒粒子组装过程中与M蛋白相互作用。ORF3蛋白是ORF3基因的产物(约25 kDa),可能促进S蛋白与宿主受体之间的相互作用。虽然ORF3不是病毒粒子的结构成分,但它保留在高尔基体中,作为离子通道发挥作用,以毒株依赖的方式增强复制和致病性。
2.1.3传播与肠道嗜性
PED在猪场主要表现为两种形式:流行性暴发和地方性感染。当PEDV传入易感猪场(大部分动物血清阴性)时,病毒迅速传播,导致所有日龄猪只近100%发病率,发生流行性PED。同时,PEDV可以持续存在导致地方性流行,主要感染失去乳汁免疫力的断奶后仔猪和新引入的血清阴性后备母猪。
PEDV靶向肠上皮细胞,通过其S蛋白与宿主受体的相互作用启动感染。虽然氨肽酶N(APN)最初被认为是主要受体,但APN基因敲除猪仍然易感,表明存在替代受体,如唾液酸、DC-SIGN、转铁蛋白受体-1和闭合蛋白,或在Vero细胞中进行体外繁殖所需的辅助因子如硫酸乙酰肝素。胰蛋白酶将S蛋白裂解为S1和S2亚基对PEDV体外生长至关重要,但一些减毒株无需此过程也能复制。这种变异性凸显了PEDV-宿主细胞相互作用的复杂性,可能反映了其从蝙蝠冠状病毒进化而来的起源。
PED在新生仔猪中最为严重,特别是血清阴性母猪所生的仔猪,高致病性毒株的死亡率接近100%。由于肠细胞坏死,临床症状(包括腹泻、脱水和绒毛萎缩)在感染后22-36小时达到高峰。较大猪只由于免疫应答更成熟及上皮细胞更新更快而抵抗力更强。
2.2免疫应答
2.2.1先天与适应性免疫
由模式识别受体(PRRs)触发的I型干扰素(IFN-α/β)是对抗PEDV关键的先天免疫应答。然而,众多体外研究表明,PEDV通过调节IFN应答和利用其结构蛋白和非结构蛋白降解免疫分子来逃避这些防御。这些逃避机制因感染PEDV毒株的毒力而异,影响临床结果和随后的适应性免疫应答。有研究表明NK细胞活性增强及IFN-γ的产生与感染仔猪病毒脱落减少和腹泻发作延迟有关。然而,这一观点受到挑战,因为PEDV会抑制NK细胞活性,并且对哺乳仔猪的IFN-γ水平影响甚微。此外,早期IFN-α分泌可能抑制哺乳仔猪后期阶段的病毒脱落,但这种效应具有毒株依赖性,非S-INDEL PEDV毒株会主动抑制IFN-α的产生。大多数研究集中在体外或新生和断奶猪的先天免疫,然而,了解妊娠母猪的这种应答对于优化疫苗设计和增强乳汁免疫力以保护新生仔猪也至关重要。
粘膜免疫,特别是由分泌型IgA(S-IgA)介导的免疫,是对抗PEDV达到长期保护的关键。病毒在肠道组织中复制诱导肠相关淋巴组织(GALT)中的抗体分泌细胞(ASCs)产生IgA和IgG抗体,这些抗体对防御至关重要。PEDV感染猪的血清IgG抗体出现较早,在感染后14-17天(dpi)达到峰值,而中和抗体在18 dpi达到峰值并持续维持高水平至42 dpi。尽管系统性抗体提供一定的保护,但粘膜应答对于持久免疫至关重要。在断奶仔猪中的研究表明,GALT和血液中较高的ASC水平,以及升高的IgG和IgA滴度,都与保护作用相关。血清、粪便和初乳中的抗S1 IgA抗体能够预测保护效果,体现了了粘膜免疫的重要性。
T细胞应答对于控制感染也至关重要。口服接种PEDV能诱导系统性CD8⁺T细胞应答,这对清除感染细胞至关重要,而以CD8⁺为主的平衡可增强免疫力。PEDV结构蛋白中保守的T细胞表位表明,利用T细胞免疫可能有助于开发广谱保护性的多价疫苗。
2.2.2粘膜免疫的作用:乳汁免疫
由于胎盘阻挡抗体转移,仔猪出生时没有母源抗体。它们依靠富含抗体和免疫细胞的初乳和乳汁提供保护,直到其免疫系统在3-6周内成熟。虽然初乳和乳汁中的抗体可以提供被动免疫,淋巴细胞也可能发挥作用,但它们在抵抗PEDV感染中的作用尚不清楚。IgG主要来源于母体血清,在初乳中占主导地位,而在乳腺局部产生的S-IgA是乳汁中的主要抗体。初乳源性免疫由系统性应答驱动,而乳汁免疫依赖于局部肠道应答。肠道中的PEDV复制刺激S-IgA浆母细胞并将免疫细胞运输至乳腺,增强了乳汁免疫。与在妊娠早期或晚期感染相比,在妊娠中期感染的后备母猪能产生更强的乳汁免疫,这强调了制备能诱导强效IgG和S-IgA应答疫苗的重要性。
PEDV作为一种肠道病原体,主要感染肠上皮细胞,需要强大的粘膜免疫应答才能有效保护。与感染呼吸道并依赖全身和气道粘膜免疫进行宿主保护的SARS-CoV-2不同,PEDV疫苗必须靶向肠道-乳腺-SIgA轴以提供持久保护。针对PEDV的粘膜免疫策略,包括减毒活疫苗和载体化平台,在刺激肠道S-IgA产生方面取得了不同程度的成功。非肠道途径的PEDV免疫能有效增加初乳IgG,但不诱导S-IgA。虽然S-IgA对粘膜保护至关重要,但血清和初乳中的高IgG水平也能降低新生仔猪死亡率。因此,最佳的PEDV疫苗应包含强效佐剂和先进的免疫原性平台以增强系统性IgG应答,同时结合刺激粘膜S-IgA产生的策略,有效靶向初乳源性和乳汁源性免疫以实现完全保护。所以,PEDV疫苗开发需要不同于SARS-CoV-2等呼吸道冠状病毒疫苗的策略,侧重于在全身应答之外诱导强大的粘膜免疫。
3、当前PEDV疫苗的挑战及SARS-CoV-2对多价疫苗开发的启示
3.1 PEDV遗传多样性及新毒株的出现
S基因的高变异性,特别是S1编码序列,导致了PEDV毒株的多样性,影响交叉免疫保护效果。PEDV毒株间S1区域的突变积累,使病毒能够逃避抗体应答并推动快速进化。相比之下,由于S2亚基主要作用是促进病毒与宿主细胞融合,该区域变化可能会破坏融合并影响病毒感染性,所以更为保守。
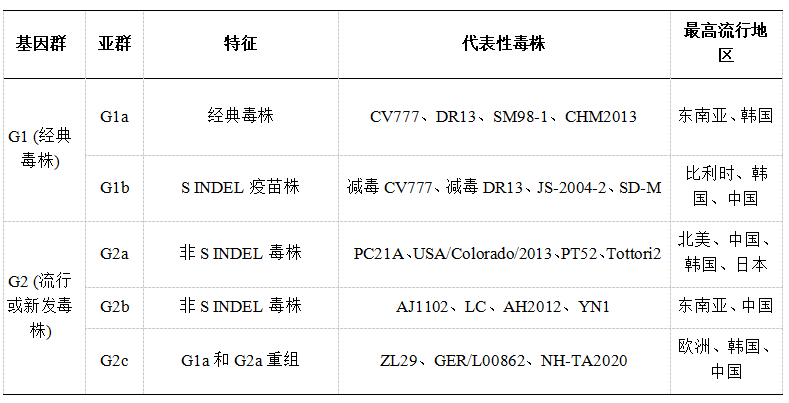
PEDV毒株分为两个基因群:经典G1毒株(1970年代在欧洲发现)和高致病性G2毒株(2010年出现于中国,此后引起全球暴发)。G2毒株的快速传播与遗传谱系间基因重组加剧了其毒力相关。这些基因群根据S基因同源性进一步细分:G1包括G1a和G1b,而G2包含G2a、G2b和G2c毒株。G1b或“S INDEL”毒株在S1亚基中存在插入和缺失,因此致病性较低。相反,高致病性“非S INDEL”毒株,特别是G2a和G2b,会导致严重暴发,而G2c毒株则是通过G1a和G2a谱系间的重组出现。从地理分布看,G1毒株在欧洲和亚洲占主导地位,而G2毒株在北美和亚洲流行(表1)。
这种遗传多样性给疫苗开发带来重大挑战,因为毒株特异性免疫无法提供跨越G1和G2毒株的保护。目前的第一代PEDV疫苗主要针对单一毒株开发,对这种遗传和抗原多样性能提供的交叉保护有限。在多种基因型共存的地区,这种局限性尤为明显,导致尽管进行了疫苗接种,疫情仍持续暴发。此外,由PEDV高突变和重组率驱动的新变种的出现,进一步削弱了疫苗效力。
因此,S蛋白仍然是疫苗设计的核心,需集中于靶向这一关键结构成分以增强交叉保护性免疫。多价疫苗策略旨在通过诱导交叉反应性免疫应答来应对这些挑战。与泛沙贝科病毒疫苗所遵循的策略类似,方法包括靶向基因型间共享的保守和免疫优势表位,以及利用创新平台,如嵌合抗原、基于纳米颗粒的疫苗和结构引导的疫苗设计。此类策略可能在地方性流行地区或高致病性变种出现期间提供持久的保护,减轻疾病暴发,并改进控制措施。
表1代表性PEDV毒株的基因群分类
3.2 S蛋白是中和抗体的主要靶点
S蛋白是病毒包膜内的同源三聚体结构,是抗PEDV中和抗体的主要靶点,它包含两个亚基:负责受体结合的S1和促进膜融合的S2。PEDV G2毒株的S1亚基由三个结构域(D0、NTD、CTD)和两个亚结构域(SD1、SD2)组成(图1)。D0结构域(PT52毒株中aa34-234)结合唾液酸,而NTD结构域辅助病毒附着,CTD作为受体结合域(RBD)与尚未鉴定的PEDV受体相互作用。S2亚基包括保守元件,如融合肽、七肽重复区(HR1和HR2)和对膜锚定和三聚体稳定性至关重要的跨膜结构域。在S1/S2连接处和S2'位点的蛋白水解裂解对膜融合至关重要,这解释了PEDV在体外繁殖时对胰蛋白酶的依赖性。由于S2的保守性及其在病毒融合中的关键作用,使其有希望成为开发广谱保护性PEDV疫苗关键靶点。
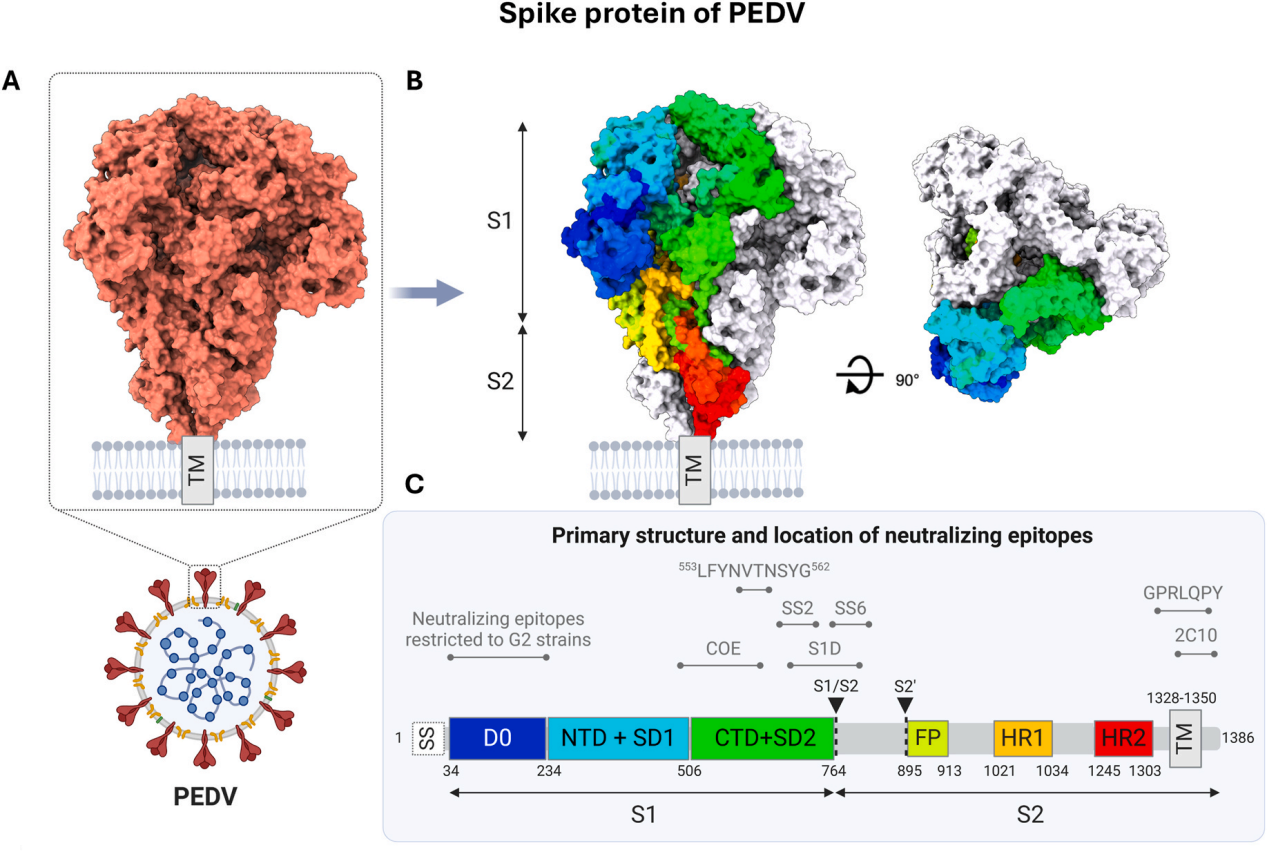
图1 PEDV S蛋白的结构与功能定位
(A)基于PT52毒株(G2)的PEDV S蛋白三聚体模型整体结构。(B)三聚体中单个单体描绘的S结构域空间排列。(C)S蛋白的一级结构,标注了中和表位的位置。SS:信号序列;D0:结构域0;NTD:S1的N端结构域;SD1:S1的亚结构域1;CTD:S1的C端结构域;SD2:S1的亚结构域2;FP:融合肽;HR1:七肽重复1;HR2:七肽重复2;TM:跨膜结构域;黑色箭头指示蛋白酶裂解位点S1/S2和S2';COE:猪传染性胃肠炎病毒(TGEV)的CO-26K等效表位。
3.2.1 S1和S2中的中和表位
PEDV S蛋白的S1和S2亚基包含多个对疫苗设计至关重要的中和表位(图1C)。S1的D0和NTD结构域含有毒株特异性中和表位,G2a毒株的D0表位是强效但交叉反应性有限的中和抗体的靶标。G1和G2毒株CTD中的其他表位被更广泛的交叉反应性中和抗体识别,包括COE表位(猪传染性胃肠炎病毒TGEV的CO-26K等效表位),该表位跨越CTD和部分SD1。经典毒株还包括位于SD2(aa 748-755)的SS2线性表位。
最近的研究鉴定出两个保守的B细胞线性表位:507FNDHSF512和553LYNNTNSYG562,它们位于S1-COE结构域内。这些表位表现出极强的免疫原性,其中553LYNNTNSYG562显示出对G1和G2基因型的强效交叉中和作用。该表位在不同PEDV毒株间的保守性强调了其作为广谱疫苗靶点的潜力。位于亚结构域SD2和S2亚基的连接处(aa 636-789)的免疫优势表位S1D,已被确定为猪抗PEDV多克隆血清中强效中和抗体的关键靶标。此外,S2中的保守线性表位,如SS2(aa 748-755)、SS6(aa 764-771)、2C10(aa 1368-1374)和GPRLQPY基序(aa 1371-1377)是关键的中和靶点。有研究表明,靶向两个亚基的IgA抗体水平与PEDV感染母猪初乳和乳汁中的中和活性相关,将来自S1和S2区域的表位纳入疫苗配方,可以在毒株特异性免疫和更广泛的保护之间取得平衡。
3.3 PEDV疫苗开发概述
3.3.1第一代PEDV疫苗:减毒和灭活病毒
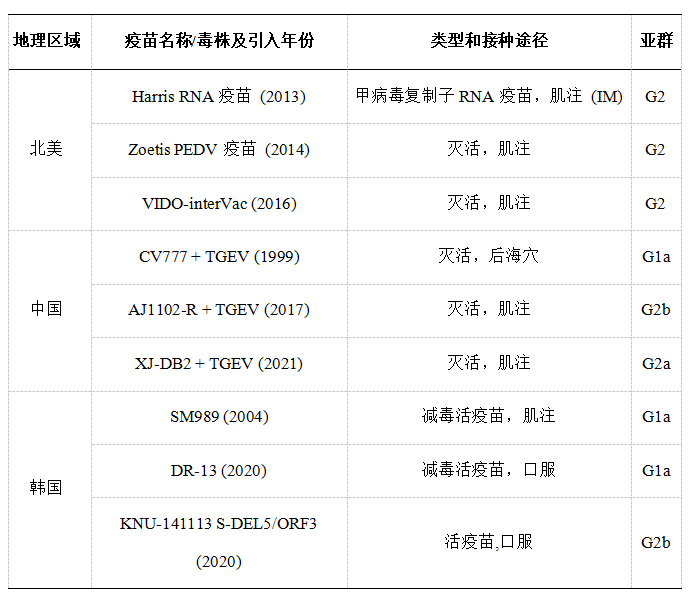
PEDV疫苗开发因地区需求和病毒动态而异。所有获得许可的PEDV疫苗都是第一代疫苗,包括减毒活疫苗和灭活疫苗(表2)。
在亚洲,基于G1a的疫苗在韩国、中国、日本和泰国广泛使用,主要以经典CV777毒株为基础,包括灭活和减毒活形式。然而,2013年高致病性G2毒株在全球的出现显著降低了其效力,促使向基于G2的疫苗转变。效力下降源于RBD中关键氨基酸的变化,特别是在亚洲G2变种中,中和表位中的七个关键替换导致PEDV基因群之间的血清中和能力降低两倍。此外,已知G2变种中的糖基化模式会影响D0和NTD结构域的分子构象。这些变化可能损害交叉保护性免疫,这可能是G1的疫苗对G2大流行毒株效力有限的原因。向基于G2疫苗的转变标志着PEDV防制方面的重大改进,尤其是在2013年暴发之后,这些疫苗现已在北美、欧洲和亚洲广泛使用。研究表明,减毒活疫苗和灭活G2疫苗在仔猪存活率上没有显著差异,两者都能有效减少病毒脱落并减轻流行毒株的影响。然而,新的PEDV变种的出现继续挑战着疫苗效力。
评估疫苗效力的一个关键限制是研究中使用的攻毒毒株多样性有限。例如,一种美国S-INDEL PEDV毒株仅能部分保护原始美国毒株,显示出抗体应答和临床结果的显著差异。同样,一种基于G2b的商业化灭活疫苗在G2b同源攻毒时减少了病毒脱落,但未能有效抵抗G1b异源毒株。此外,一种S-INDEL PEDV疫苗降低了仔猪死亡率(未接种后备母猪为74.4%,接种组为36.4%),但窝间差异较大,强调了需要更有效的疫苗诱导粘膜免疫。此外,减毒活疫苗存在病毒重组风险,可能导致毒力更强的毒株出现。研究开发靶向PEDV基因群间保守表位的新疫苗配方以提高疫苗效力并确保能够持续地控制PEDV至关重要。
表2北美、中国和韩国代表性获批或有条件获批的PEDV疫苗
3.3.2新一代疫苗平台
为应对新出现的PEDV G2毒株,研究人员利用新型平台开发了新一代疫苗,包括亚单位疫苗、纳米颗粒、病毒样颗粒(VLPs)和基于核酸的疫苗(mRNA、DNA、病毒载体)。每个平台都具有独特的优势,包括安全性、快速适应性和靶向免疫应答,但其效力在不同研究中存在差异,表3概述了第一代和新一代平台的比较。
亚单位疫苗使用纯化的抗原,如全长S、S1或S2蛋白,结合佐剂以减轻与活疫苗或灭活疫苗相关的生物安全风险。虽然像大肠杆菌和昆虫细胞这样的平台已用于抗原生产,但哺乳动物细胞因其具有对免疫原性至关重要的支持正确糖基化和维持蛋白质构象的能力而更受青睐。已测试了多种PEDV亚单位疫苗构型,包括全长S蛋白、S蛋白三聚体、S1或S2亚基以及特定的中和表位。
纳米颗粒和VLPs因其最佳尺寸(20–200 nm)和重复的表面结构而成为高效的疫苗平台,可增强抗原摄取并刺激强烈的免疫应答。这些平台要么将抗原封装在脂质核心中(如mRNA疫苗),要么通过化学偶联或基因融合将抗原展示在其表面。基于纳米颗粒的PEDV疫苗研发已整合了多种抗原,包括S蛋白亚基、特定的中和表位以及N或E蛋白以拓宽免疫保护。
核酸疫苗将编码抗原的遗传物质直接递送到宿主细胞中,诱导内源性蛋白合成,模拟自然感染并激活先天性和适应性免疫应答。mRNA疫苗存在冷链储存要求等限制,具有适应性强和生产快速的优势。除mRNA疫苗外,研究者还探索了各种细菌和病毒载体用于递送PEDV抗原。细菌载体,如乳酸杆菌属(Lactobacillus spp.)和沙门氏菌属(Salmonella spp.),已被用作PEDV抗原的粘膜递送载体。同样,病毒载体如腺病毒、副痘病毒、杆状病毒和伪狂犬病毒已被研究作为PEDV疫苗平台。SARS-CoV-2疫苗开发的进展为合理的抗原设计和多价PEDV疫苗开发提供了宝贵的经验。
表3 PEDV多毒株疫苗开发的第一代和新一代平台比较

3.3.3 SARS-CoV-2对多价PEDV疫苗开发的启示
虽然新一代平台已被探索用于单毒株PEDV疫苗,但对多毒株PEDV疫苗的研究仍然有限。本节回顾了关于SARS-CoV-2泛沙贝科病毒疫苗和PEDV研究,以强调尖端抗原设计策略如何应用于各种平台。
抗原设计的进步,特别是基于结构和嵌合的方法,在开发针对抗原可变病原体的广谱保护性疫苗方面至关重要。例如,基于结构的疫苗设计可以稳定PEDV刺突蛋白的前融合构象,增强免疫原性。此外,结合PEDV基因群间保守表位的嵌合抗原可能促进更广泛的交叉保护。除了抗原设计,尖端平台如mRNA、纳米颗粒和病毒载体提供了多功能递送系统,可增强免疫应答效力并实现快速的抗原定制。这些技术已被证明对SARS-CoV-2有效,可以灵活整合来自不同PEDV基因群的保守和可变抗原,从而支持多毒株PEDV疫苗的开发。
几种泛沙贝科病毒疫苗正在进行临床试验,提供了可适用于PEDV疫苗研发实例。例如,沃尔特·里德陆军研究所的刺突铁蛋白纳米颗粒(SpFR)疫苗展示了纳米颗粒平台如何以多价形式呈递抗原来引发广谱的免疫。同样,VBI疫苗公司的三价eVLP疫苗(VBI-2901)包含了来自多种冠状病毒(SARS-CoV-1、SARS-CoV-2和MERS-CoV)的S蛋白,显示了多价方法应对抗原多样性的潜力。SK Biosciences的RBD-纳米颗粒(GBP511)包含了多样化的RBD,突显了呈递保守和可变表位以引发交叉反应性免疫应答的功效。这些方法可以通过靶向保守表位(如S2亚基)以及可变区域来将交叉保护潜力最大化,从而适用于PEDV疫苗。
总之,这些SARS-CoV-2策略为解决PEDV疫苗学中抗原多样性的挑战提供了一个强有力的框架(图2)。
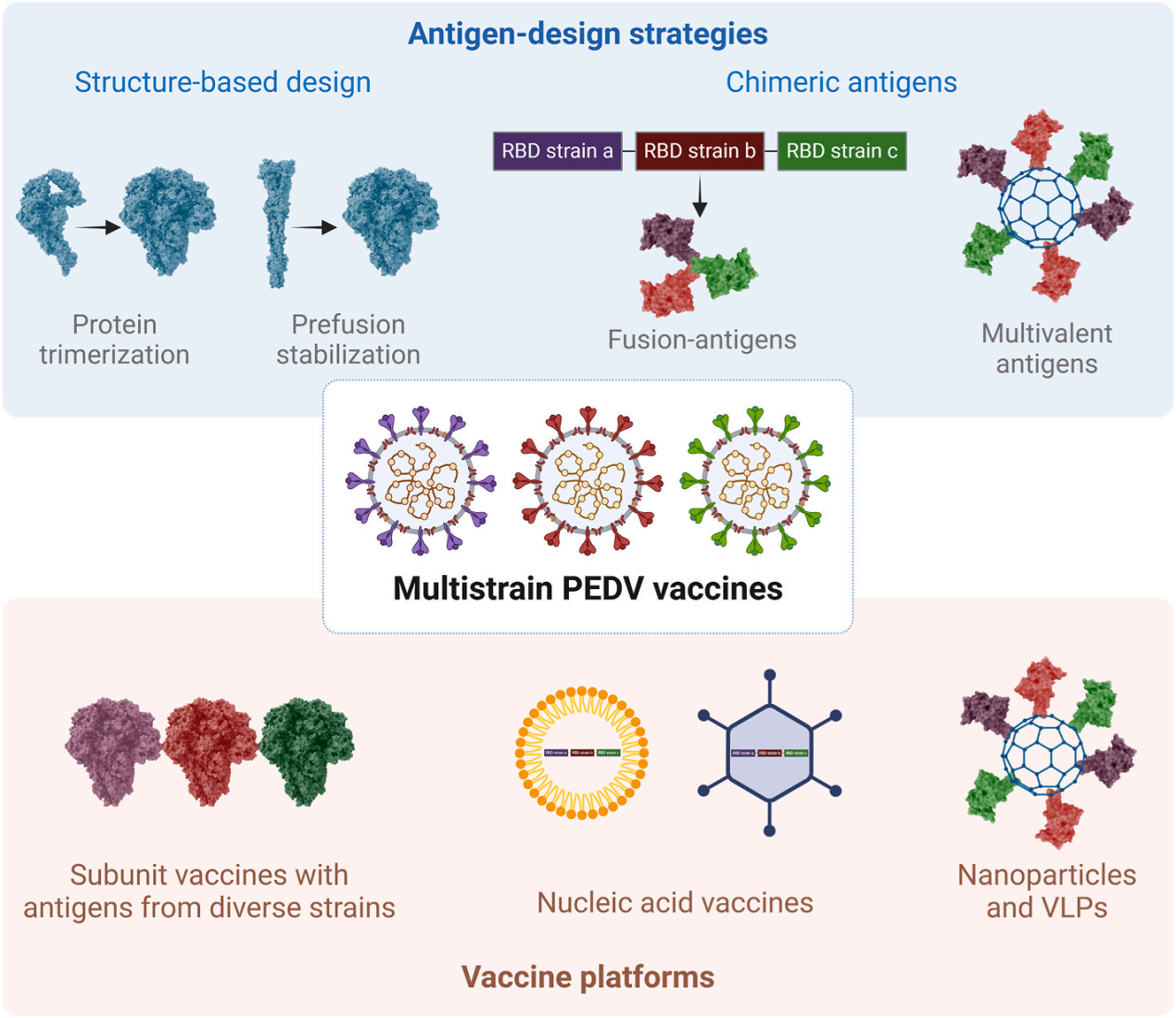
图2多价PEDV疫苗研发方案示意图
通过抗原设计和疫苗平台构建多价PEDV疫苗的示意图。基于结构的设计方法(如蛋白质三聚化和预融合稳定化)可保持PEDV S蛋白的天然构象,从而增强免疫识别与稳定性。嵌合抗原设计方案包括融合多毒株抗原或整合保守抗原(如N蛋白),以及通过多价抗原呈现保守表位以扩大免疫保护范围。这些抗原可通过多种载体递送,包括含有不同PEDV毒株抗原的亚单位疫苗、异源初免-加强方案、核酸疫苗(mRNA和DNA),以及纳米颗粒或类病毒颗粒(VLPs)。晶体结构(PDB编号:7W6M)。
4、广谱保护的抗原设计方案
4.1基于结构的抗原设计和保守抗原
基于结构的抗原设计旨在通过将抗原稳定于可高效刺激中和抗体产生的特定构象中,从而增强免疫识别效能。表位稳定化、三聚化基序嵌入、二硫键工程及点突变等技术,可有效将抗原锁定于免疫优势构象中。在SARS-CoV-2疫苗研发中,三聚化基序与前融合稳定化策略是此类设计的典型范例:三聚化模拟了刺突蛋白(S蛋白)的天然三聚体空间结构,而前融合稳定化则将其构象锁定于融合前状态,最大限度保留关键中和表位的可及性。这些策略显著增强了对毒株间保守表位的抗体应答广度,拓宽了整体免疫反应谱。其在SARS-CoV-2疫苗中的成功应用,为开发覆盖多毒株的PEDV疫苗提供了重要技术范式。
4.1.1用于增强S蛋白稳定性和免疫原性的三聚化基序
引入三聚化基序,如T4纤连蛋白(foldon)结构域,已被证明是增强疫苗抗原稳定性和免疫原性的有效方式。以泛β冠状病毒亚单位疫苗Om-S-MERS-RBD为例:通过T4 foldon基序稳定化的抗原,在BALB/c小鼠模型中维持了三聚体稳定性并提升了抗原性,成功诱导产针对MERS-CoV、SARS-CoV和SARS-CoV-2的交叉中和抗体应答。
早期的PEDV疫苗研究主要聚焦于不同表达系统中生产的G2a毒株的可溶性S1结构域。虽然这些疫苗可诱导部分保护性免疫并缓解临床症状,但在控制病毒排放和腹泻方面效果较差。为优化免疫效果,研究者将三聚化基序整合到全长S或S1亚基中。例如,在S1亚基上融合鸡软骨基质蛋白三聚化基序,使小鼠模型的IgG和中和抗体(nAb)应答显著增强。
基于上述策略,研究者开发了一种双价亚单位疫苗,该疫苗整合了经T4纤连蛋白结构域稳定化的PEDV G2a和G2b毒株的全长S蛋白。在观察到仔猪接种灭活G2a或G2b PEDV疫苗后交叉保护有限后,作者采用1:1抗原比例配制了双价亚单位疫苗,并用M103佐剂增强免疫原性。在主动免疫试验中,仔猪经基础免疫和加强免疫后,显示出强效的中和抗体应答、临床症状减轻以及攻毒后病毒载量降低。被动免疫结果显示,妊娠母猪免疫后通过初乳传递母源抗体使仔猪抵抗力增强、临床症状减轻。这些结果凸显了双价、三聚化亚单位疫苗作为高效广谱PEDV保护候选疫苗的技术优势。
4.1.2 S蛋白的前融合稳定化
除了三聚化,稳定全长S蛋白中S2亚基的前融合构象可以进一步增强nAb应答。与SARS-CoV-2类似,PEDV S蛋白易于从前融合状态转变为后融合状态,损害关键的中和表位。这种转变是由受体结合后宿主蛋白酶(如胰蛋白酶)在S2'位点切割触发,进而暴露融合肽并促进其插入宿主细胞膜,启动膜融合过程。S2亚基的结构动力学及其在构象重排中的关键作用,凸显了稳定该区域以保持表位完整性的重要性。在SARS-CoV-2疫苗中已成功应用前膜融合稳定化技术,如在S2中进行脯氨酸替代和引入二硫键,可将S蛋白锁定于前融合状态,保持表位完整性并提高生产过程中的抗原产量。
然而,前融合稳定化在PEDV疫苗中尚未得到广泛研究,目前仅有一项报道的研究评估了在昆虫细胞中表达,并在小鼠和仔猪模型中评估的前融合稳定化全长S蛋白。经CpG5和MF59联合佐剂配伍的稳定化S蛋白,在小鼠体内诱导了强烈的Th1型免疫应答,表达为IgG2a和IFN-γ,同时在仔猪中显著增强了中和抗体滴度和CD8⁺T细胞应答。但该研究未设置攻毒实验组及非稳定化S蛋白对照组。未来的研究应探索采用哺乳动物表达系统以保留天然糖基化模式和正确折叠,进而优化PEDV疫苗的免疫原性。
4.1.3改造的S2作为保守的亚单位抗原
尽管大多数中和抗体靶向S1亚基,但S2亚基在PEDV、SARS-CoV-2和其他冠状病毒中高度保守,使其成为多价和泛沙贝科病毒疫苗开发的潜力靶点。然而,在SARS-CoV-2和其他β冠状病毒中,虽然部分靶向S2的抗体显示出广谱反应性,但其由浆细胞和记忆B细胞产生的水平远低于靶向S1的抗体,从而限制了其整体中和效力。为克服这一限制,需要基于结构的抗原设计,以刺激接种宿主产生高水平的广谱反应性S2抗体。
在SARS-CoV-2研究中,已开发了几种仅含S2的免疫原(即“仅茎区(stem-only)”免疫原),其中S2亚基经前融合稳定化改造。这种设计旨在诱导阻断膜融合过程的抗体,从而实现有效中和病毒。然而,在缺乏S1的情况下表达处于前融合构象的S2亚基仍具有挑战性。两项研究通过引入二硫键和脯氨酸突变,成功设计了前融合稳定的S2三聚体,显著稳定性和免疫原性。免疫小鼠产生了对同源和异源SARS-CoV-2毒株均有效的广谱中和抗体。类似地,一种稳定的MERS-CoV S2抗原在小鼠中引发了对MERS-CoV和SARS-CoV-2的交叉反应性抗体。
另一种方法聚焦于模拟融合中间状态的七肽重复1(HR1)结构域,在大肠杆菌中创建HR121亚单位疫苗,包含HR2两侧为两个HR1的序列。这种免疫原诱导了交叉中和抗体,并在hACE2小鼠、仓鼠和恒河猴中提供了针对多种SARS-CoV-2变种的近乎完全保护。一种创新型性策略使用了靶向S2结构域的“低糖(low-sugar)”mRNA疫苗,通过删除糖基化位点暴露保守表位。免疫小鼠表现出强烈的CD8⁺T细胞活化,以及对SARS-CoV-2变种以乃至其他冠状病毒(如MERS和SARS)的交叉中和抗体,表明其具有广谱、抗变异保护的潜力。
对于PEDV,研究人员开发了一种靶向G2a毒株S蛋白的HR1和HR2结构域的纳米颗粒疫苗。将HR蛋白与幽门螺杆菌铁蛋白融合,可自组装成纳米颗粒,在该疫苗小鼠模型中诱导了PEDV特异性的IgG和中和抗体。
4.2嵌合抗原
嵌合抗原设计将不同的表位、蛋白结构域或整个蛋白组合成单一多肽链,以增强免疫原性并刺激更广的免疫应答。这种方法成功用于泛沙贝科病毒疫苗研发,并有望用于PEDV疫苗。包括展示来自多种SARS-CoV-2毒株的前融合稳定化S蛋白的纳米颗粒、来自多个变异株的RBDs,或包含S或S1和N蛋白的融合构建体。这些嵌合构建体可以作为可溶性蛋白递送,或通过mRNA平台或病毒载体递送。
4.2.1包含RBD/COE的嵌合抗原
研究人员开发了一种基于三聚体RBD的可溶性蛋白疫苗,通过将Omicron RBD与来自其他变异株的两个具有关键突变的改造型RBD相结合,实现对SARS-CoV-2变体的广泛中和能力。该疫苗在哺乳动物细胞中表达,并在大鼠模型中评估。结果显示,与灭活疫苗和仅包含Omicron RBD的同源三聚体RBD构建体相比,该疫苗能够诱导更优越的交叉中和抗体应答。另一个引人注目的设计称为“列车模型”,将武汉株S1骨架与来自Omicron、Delta和Beta变异株的RBD相结合。在哺乳动物细胞中表达并用辅以MF59类佐剂进行免疫。在叙利亚仓鼠模型中,该疫苗诱导了广泛的免疫应答,并对多种变异株具有保护效力。此外,该疫苗在2–8°C下可以稳定保存超过一年,这一特性为其在资源匮乏环境中的应用提供了重要支持。
在推进多价疫苗过程中,研究者设计了“四聚体纳米笼”,用于在单个纳米颗粒上使用SpyTag/SpyCatcher系统展示来自四种沙贝科病毒(SARS-CoV-2、SHC014、Rs4081、RaTG13)的RBD。这些纳米笼在BALB/c小鼠中诱导了对所对应的的和错配的沙贝科病毒的中和抗体滴度,充分显示了其作为广谱疫苗平台的潜力。
在PEDV研究中,一种靶向G2a毒株的纳米颗粒疫苗利用了基于铁蛋白的自组装纳米颗粒。该疫苗用SpyTag/SpyCatcher技术整合了来自PEDV S1蛋白的NTD、CTD或NTD/CTD组合。其中,组合的NTD/CTD纳米颗粒在小鼠中引发了最强的IgG和中和抗体应答,并且通过初乳传递的母源抗体为仔猪提供了保护,减轻了临床症状及减少了病毒载量。这项研究证明了在纳米颗粒疫苗中结合S1-NTD和S1-CTD具有增强PEDV效力的潜力。
PEDV疫苗最新研发进展包括mRNA疫苗,mRNA编码未修饰的全长S蛋白或整合了NTD、COE表位及S2区域中和表位的多表位嵌合S。这些疫苗由G2b毒株合成并封装在脂质纳米颗粒(LNPs)中。在小鼠模型中,S mRNA疫苗的体液和细胞免疫应答优于多表位嵌合S mRNA疫苗,表现为更高的中和抗体和T细胞活化水平。在仔猪模型中,免疫S mRNA疫苗使攻毒仔猪临床症状及肠道损伤减轻、病毒载量降低。通过母源抗体的被动免疫进一步显示了mRNA平台在疫苗快速开发中的重要作用。
4.2.2 S+N融合抗原
虽然S蛋白是SARS-CoV-2和PEDV亚单位疫苗中中和抗体的主要靶点,但N蛋白由于其跨PEDV毒株和SARS-CoV-2变异株的高度保守性(90-94%氨基酸相似性)及其在病毒组装和基因组包装中重要作用,有潜力成为T细胞介导免疫的靶点。虽然N蛋白在T细胞介导的PEDV防御中的作用尚不清楚,但对SARS-CoV-2的研究已证明其能够引发与病毒控制相关的持久T细胞应答。在SARS-CoV-1感染后长达17年仍能检测到N蛋白特异性记忆T细胞,表明N蛋白能引起广泛持久T细胞免疫。因此,将N蛋白整合到PEDV疫苗中可以延长T细胞应答,加强S蛋白诱导的抗体应答。
一种前景广阔的疫苗策略涉及嵌合亚单位疫苗,该疫苗将S蛋白或其亚基(S1或S2)与N蛋白结合,以增强免疫应答。在SARS-CoV-2疫苗的研究中,一种编码前融合稳定化S和N蛋白的双mRNA疫苗,在BALB/c小鼠和叙利亚仓鼠模型中进行了测试,评估其对Delta、Omicron BA.1、BA.5和BQ.1变异株的效力。这种S+N组合策略显著降低了肺病毒载量,防止了体重减轻,并且即使在没有检测到针对进化分支较远的亚变体的中和抗体的情况下,其保护效果也优于仅包含S蛋白的疫苗。此外,肺组织中强烈的S蛋白特异性和N蛋白特异性CD8⁺T细胞应答,进一步凸显了S+N组合在诱导广泛T细胞免疫方面的潜力。
采用病毒载体疫苗和纳米颗粒递送平台也观察到了类似结果。例如,鼻腔给药的NanoSTING-SN多抗原疫苗(包含S和N蛋白),在动物模型中不仅成功阻断了SARS-CoV-2传播,还诱导了强烈的体液免疫和细胞免疫。虽然目前尚未开发出采用S+N嵌合策略的PEDV疫苗,但借鉴此类方法有望应对PEDV的遗传多样性挑战,并提升疫苗效力。
4.3混合多毒株抗原与异源初免-加强策略
通过用混合多毒株抗原免疫怀孕母猪或实施异源初免-加强策略,可以实现针对多种PEDV毒株的广泛保护。混合多毒株抗原结合了来自多个病毒毒株的表位或亚单位蛋白,从而通过靶向保守和毒株特异性表位来拓宽免疫覆盖范围。异源初免-加强方案通过按顺序免疫不同平台(如亚单位、病毒载体、mRNA和灭活疫苗)的疫苗来增强这种效果,以优化体液和细胞免疫应答,同时促进持久免疫。
异源疫苗接种的优势在SARS-CoV-2研究中得到充分证明。例如,在人类患者中,相较于同源方案,将腺病毒载体疫苗与mRNA疫苗(BNT162b2或mRNA-1273)联合使用,显著提高了S蛋白特异性IgG、CD8⁺T细胞活化和中和抗体水平,同时保持了一定的反应原性。同样,在小鼠模型的临床前试验中,编码变体株特异性S蛋白的异源LNP-mRNA方案可诱导比同源野生型疫苗方案更强的中和效价、更广泛的T细胞应答和增强的多功能性CD8⁺T细胞活化。
针对PEDV的专项研究与这些发现相吻合。一种整合了PEDV G2a和G2b毒株的三聚化S蛋白混合物的双价亚单位疫苗,相比单价疫苗展示出更优的免疫原性,不仅诱导了更高的中和抗体滴度、IgG和IgA应答水平,还能提供更好的攻毒保护。接种疫苗的仔猪腹泻评分更低、病毒载量减少、肠道损伤减小,而且接种疫苗的母猪的初乳能为子代提供被动免疫。
尽管仍需深入研究,但将多毒株抗原混合与异源初免-加强策略相结合,利用抗原多样性并优化免疫应答,在开发具有广谱保护性的猪流行性腹泻病毒(PEDV)疫苗方面仍前景广阔。
5、结论与未来方向
研发针对PEDV的多价疫苗迫在眉睫,但目前研究有限。PEDV频繁的突变和重组事件对设计能引起持久和全面免疫反应的疫苗提出了重大挑战。尽管现有疫苗降低了死亡率,但常无法阻止病毒排放,亦不能有效应对新兴变异株,这凸显了对创新疫苗解决方案的需求。
鉴于猪流行性腹泻病毒(PEDV)对全球养猪业造成的严重经济损失和健康危害,开发具有广泛交叉保护效力的疫苗对于控制PED暴发至关重要。新生仔猪的高死亡率及其导致的重大经济损失,突显了有效疫苗的迫切需求。缺乏此类解决方案将加剧生产者的经济负担,进一步威胁粮食安全。
应对这些挑战,亟需整合来自SARS-CoV-2疫苗研发的成熟策略,例如先进的抗原设计和新一代疫苗平台。确保新疫苗能够以成本效益高的方式生产和交付,对于其在资源有限地区的可及性和可扩展性至关重要。兽医疫苗平台的设计不仅要优先考虑免疫原性,还需兼顾生产效率和制剂稳定性。因此,提升mRNA疫苗的稳定性与可扩展性,同时积极探索亚单位疫苗、病毒载体疫苗及灭活病毒疫苗等平台技术,对于实现广泛采用依然至关重要。
考虑到PEDV的肠道嗜性,粘膜免疫提供了另一极具前景的途径,能够激发强大粘膜免疫和全身免疫应答的疫苗,可显著减少病毒脱落和传播。开发结合粘膜递送系统与能增强母猪乳汁免疫力的佐剂是提升疫苗效力的重要途径。此外,开发能够区分感染动物与免疫动物(DIVA)的疫苗,将有助于改善疫情监测并实施更有效的防控措施。
总之,开发有效的多价PEDV疫苗,需要采取整合先进抗原设计、创新疫苗平台及经济考量的多学科策略。通过借鉴SARS-CoV-2疫苗开发的经验教训并促进多方合作,养猪业有望找到控制PEDV并减轻其影响的可行解决方案。
注:以上数据仅供参考,不作任何投资建议





