2011年以来新一轮猪伪狂犬病爆发流行,给我国养猪业造成了巨大经济损失。经研究新型伪狂犬病毒多个基因碱基发生了标志性突变和连续性缺失,毒力明显增强。大量的研究报道表明,伪狂犬病毒在我国各省区广泛存在,gE抗体阳性率依然很高,猪场伪狂犬控制和净化面临较大的压力。
1 经典伪狂犬病流行历史
1813年美国首次临床上描述了伪狂犬病毒对牛感染发病,病牛表现为极度瘙痒,最后死亡。1849年瑞士最先用伪狂犬对该病进行了命名。1902年,匈牙利内科医生Aujeszky报道了在患病公牛、犬和猫上分离到感染源,并证明这种病原与狂犬病毒不同,始称Aujeszky`s disease(AD)。1910年SchmiedhofferF发现致病因子是病毒,称为pseudorabies virus(PrV)。1933年德国Erich Traub用兔脑、睾丸组织体外培养PrV获得成功。1934年Sabin and Wright将PrV划归疱疹病毒群。1995年ICTV第六次报告列入疱疹病毒可α-疱疹病毒亚科水痘病毒属,分类学名猪疱疹病毒ǀ型。目前,该病主要流行于欧洲东部和东南部、拉丁美洲、非洲和亚洲;欧洲大部分国家,如德国、荷兰、丹麦、瑞典、匈牙利、英国等,以及加拿大、美国、新西兰等国家和地区已在饲养环境的猪群中消灭了伪狂犬病。
我国1947年首次报道伪狂犬病。20世纪90年代,我国伪狂犬病广泛发生,给养猪业造成严重经济损失。2000-2011年,随着伪狂犬病疫苗广泛使用,猪伪狂犬病的防控已经做得相当不错,临床上很少有猪场发病,很多规模化猪场伪狂犬野毒感染己经净化为阴性场,该病曾经是我国最有希望达到净化标准的猪病。
2 新型伪狂犬病流行情况
2.1 流行情况
2011年下半年起,我国爆发了新一轮新型伪狂犬病疫情,很多伪狂犬gE阴性场,gE抗体快速转阳,伪狂犬疫苗免疫群也发生典型的伪狂犬疫情。据佛山科学技术学院白挨泉教授报道,2011年11月份首先在天津等地区开始发现生长育肥猪表现神经症状而死亡的新型伪狂犬病现象;2012年华北、华中、华东等地区表现出同样的伪狂犬病疫情;2013年疫情开始扩散到南方多省,以广东、广西等地区表现明显;2014年福建、山西、云南等地区猪场也出现新型伪狂犬病毒感染现象;目前,我国主要养猪区域都有本病发生的报道。新型猪伪狂犬病感染的主要表现,在很多伪狂犬病阴性场gE抗体检测突然转阳,阳性场比例暴增,猪群突然出现发烧咳嗽,采食量减少。新型猪PRV流行株与经典毒株相比对猪的致病性更强,很多猪场首先发病的是生长育肥猪,随后出现母猪流产、产死胎、产弱仔以及哺乳仔猪、保育仔猪、生长育肥猪均有可能出现腹泻、呼吸道症状、神经症状等,病亡率很高。
Tan等(2021年)对108篇PRV血清学检测相关报道进行了汇总分析,2011年至2020年上半年期间,从中国29个省份共检测256326份样品,gE抗体阳性76553份 ,阳性率29.87%。
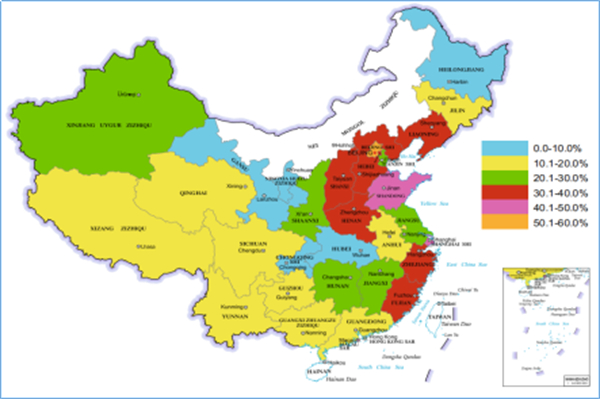
图1:2011-2020年不同地区伪狂犬gE抗体检测结果汇总分析图
据华中农大诊断中心的检测结果,2012年-2019年猪伪狂犬病病毒(PRV)在国内猪场的病原检出率和gE抗体阳性率均呈现先上升后下降的趋势,但是在2020年猪伪狂犬病gE抗体阳性率出现了明显的反弹。
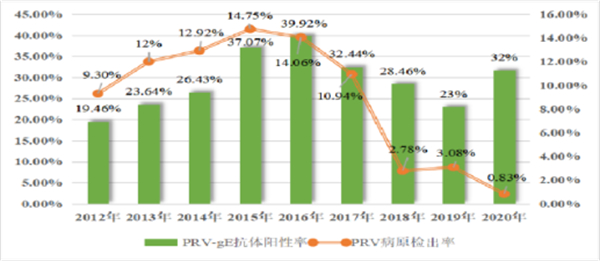
图2 2012-2020年猪场PRV和gE抗体检出阳性率
袁淑萍(河南省疫控中心)等对河南1384个场次的41755份猪血清进行gE抗体检测,2017-2019年不同年份猪群gE抗体个体阳性率分别为26.21%、28.82%和25.31%,场群阳性率分别为53.86%、52.09%和51.50%。
从各地伪狂犬流行病学调查报告显示,中国各省份都存在伪狂犬病毒流行,但流行程度不一,多为潜伏感染。不同猪场野毒感染程度差异大,不同生长阶段伪狂犬病毒感染率也不同,主要呈散发和地方性流行。总体来看,gE抗体阳性率仍处于较高水平,野毒依然广泛存在,一旦忽视了伪狂犬的免疫防控,容易造成猪场伪狂犬不稳定,我国规模种猪场伪狂犬病的全面净化仍面临巨大挑战。
2.2 变异伪狂犬病毒的分子特征
通过基因扩增测序,新分离的PRV出现基因变异,主要在TK,gB,gC和gE发生多个碱基的标志性突变和连续性缺失:与Bartha株相比,毒力相关基因RR基因第31位至36位出现6个连续碱基(第11位和第12位氨基酸)的缺失;与Bartha株比较,变异伪狂犬病毒gB基因的第224-232位发生9个碱基GCCCCGGCC的缺失,也就是3个AA缺失。变异株伪狂犬gE 蛋白第48位和第492位各有1个天冬氨酸的插入,虽然部分经典株在第48位也有一个天冬氨酸的插入,但变异株在此位点却高度一致(赵鸿远等,2014年)。变异株伪狂犬病毒gC基因63-69位连续7个氨基酸AAASTPA的插入(叶超等,2015年)。
哈兽研研究团队首次提出PRV存在两个基因型(G1、G2),我国流行的为基因2型。基于gB、gC、gD 、gE等基因的进化树分析结果显示,欧美分离株可归为一大分支,为基因Ⅰ型,中国分离株与韩国分离株可归为另一大分支,为基因Ⅱ型;而基因Ⅱ型又分为2个小分支,2010年前分离的毒株与MinA株和Ea 株为代表的亲缘关系较近,处于同一小的分支中,为经典毒株;2012年以后分离的流行毒株亲缘关系较近,与以MinA株和Ea 株为代表的亲缘关系较远,处于另一分支中,为伪狂犬变异毒株。
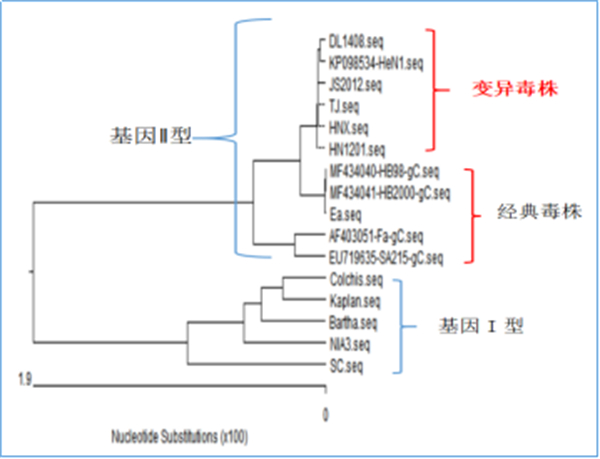
图3 伪狂犬gC基因序列分析
2.3 新型伪狂犬爆发流行的原因分析
造成我国2011年以来新型伪狂犬病流行的原因是多方面的:
(1)伪狂犬病毒发生了变异,毒力明显增强。
(2)经典毒株疫苗产生保护力下降,原有效价标准的疫苗所产生保护力不足以抵抗强毒的入侵。
(3)免疫程序不合理,免疫频次不够。
(4)猪群gE阳性率居高不下,环境病毒污染面广,稍有不慎就有可能造成猪群波动。
(5)免疫抑制因素广泛存在加重疫情严重程度。

针对以上原因,要想提高猪伪狂犬病疫苗的保护率可从以下几个方面入手:
(1)提高疫苗效价,经典毒株效价一般要≥106.3TCID50才有更好的保护。
(2)适当提高伪狂犬疫苗的免疫频次,生产种猪1年普免4次;仔猪1-2日龄喷鼻免疫,40-50日龄肌注免疫,80日龄左右加强免疫;后备猪配种前免疫2次。
(3)加强生物安全管理和生产流程管理。
(4)加快变异毒株伪狂犬病疫苗的研究和应用。变异毒株疫苗具有更强的针对性和更高的保护效果,是猪场伪狂犬病净化的首选。华派生物集团和哈尔滨兽医研究所田志军研究团队合作的变异伪狂犬5基因缺失疫苗(TP株)的研制工作进展顺利,预计明年正式上市。